Od Ole Pedersen, Claus Christensen i Troels Andersena
Slabiji rast biljaka u akvarijumima najčeöće se pripisuju neadekvatnom izvoru svetla iznad akvarijuma i ukoliko se konsultujete sa nekim ko zna viöe od vas, savet je uvijek bio da se poveća intezitet svetla prije nego öto se bilo öta drugo uradi. Novija istraûivanja pokazuju da to ne mora uvek da bude ispravan savet, ovo se posebno odnosi na akvarijume u kojima se ne dodaje CO2.
U knjigama vezano za akvaristiku, joö uvijek moûete pronaći tekst koji se odnosi na to da je dovoljno da vam jedan od faktora koji su bitni za rast biljaka nije adekvatan i da na taj način ne budete zadovoljni sa rezultatom. Ovo činjenjično stanje je poznato kao princip Liebig.
Slika 1. Na slici je prikazana ilustracija Liebigovog principa. U konkretnom primeru Bor limitira rast biljaka öto je ilustrovano isticanjem vode iz bureta kada se dostigne granica u rastu biljaka.
Justus Liebig je poznati nemački hemičar (12 May 1803 ñ 18 April 1873) koji je radio većim delom radnog veka na prihrani biljaka. Justus Liebig je postavio tezu da je dovoljan jedan i samo jedan faktor koji moûe da ograniči rast biljaka na neko vreme. I danas je nejasno da li je sam Liebig razvio ovu tezu, slika 1 jednostavna staklena cev, koja je delomično ispunjena vodom, uveliko je pridonela odrûavanju ove teze o nedostatku resursa. Nekoliko decenija kasnije je ustanovljeno da nedostatak jednog činioca neće uticati znatno na razvoj kopnenih biljaka, takvo razmiöljanje o ograničenju resursa se zadrûalo i na okvirima podvodnih biljaka u narednih dvadeset godina. U novijem periodu nekoliko studija vezane na ovu temu su pokazale da je uticaj povezanosti izvora svetlosti i CO2 proces poznat kao fotosinteze ima znatan uticaj na rast biljaka (Maberly 1985, Madsen and Sand-Jensen 1994). U ovom tekstu biće prikazani rezultati iz eksperimenata gde se eksperimentisalo sa ograničenim koncentracijama CO2 i slabim izvorom svetla, koji su u prirodi dva glavna ograničavajuća faktora za rast podvodnih biljaka.
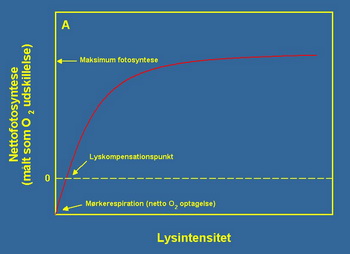
Da bi razumeli kako biljke reagiraju na upadni zrak svetlosti, izvorno svetlo koje je koriöteno na podvodnim biljkama öematski je prikazano na slici 2A. Eksperiment je pokazao da ukoliko imamo vrlo nizak intenzitet izvora svetla ono nije dovoljno za odrûavanje normalnih procesa fotosinteze i kao takvo nije zadovoljavajuće.
Drugim rečima, kod biljaka preovladava proces disanja umesto procesa fotosinteze. Međutim na određenom nivou svetla, ova dva procesa se izjednačavaju i tada dobijamo definiranu tačku koja odgovara biljkama. Ukoliko povećamo intezitet svetla, biljke će u tom slučaju biti stimulisane za proces fotosinteze. Na izuzetno visokom nivou izvora svetla, kao rezultat imamo izlazak iz procesa fotosinteze sve do momenta kada se proces fotosinteze svodi na nivo kao kada je proces fotosinteze bio najintezivniji. Sa te tačke povećanje izvora svetla neće imati uticaj na stimulisanje procesa fotosinteze.
U prirodnim uslovima, rast vodenih biljaka često je uslovljen od strane ograničene dostupnosti svetla. Biljke prirodnu svetlost efikasno apsorbuju kroz vodu, jedan deo svetlosne energija se pretvara u toplotnu energiju, ukoliko voda sadrûi rastvorena organska jedinjenja - na primer huminska kiselina u tamnijim vodama - apsorpcija svetlosti je efikasnija. Samo apsorbovanje sunčeve svetlosti i rastvorenih organskih jedinjenja sa povećanjem vodenog stuba opada transparentnost vode i samim time opada iskoriötavanje od strane vodenih biljaka öto dovodi do propadanja istih, u ovom slučaju na ovakvim staniötima samo plutajuće biljke mogu da opstaju. Iz svega gore navedenoga da se izvući zaključak koliko je sunčeva svetlost bitan parametar za biljke.
Vekovnim evoluiranjem podvodne biljke su same razvile vrlo uspeöan način lakog iskoriöćenja slabog izvora svetlosti. Ukoliko biljka ima dovoljno rastvorenih organskih jedinjenja na raspolaganju, sama biljka će veći deo svoje energije da uloûi na iskoriötavanje sunčeve energije uz pomoću pigmenata karotenoida, ksanthophylls i joö vaûnijeg, hlorofila.
Hlorofil je u suötini zeleni pigment koji apsorbuje svetlo i pretvara ga u hemisku energiju, koja se moûe koristiti za rast i razvoj ćelija. Na ovaj način biljka se osigurava da dostigne svojim rastom öto adekvatniju poziciju da bi izvor svetla i samim tim dobijena energija u procesu fotosinteze bila öto efikasnije iskoriötena a ne samo da se prenosi kroz biljku. Takođe je neophodno da biljak sadrûi puno hlorofila da bi dostigla maksimalno iskoriötenje u procesu fotosinteze, ali velika količina hlorofila opet nema velike koristi ako energija koja se dobija ne moûe biti iskoriötena za neorganski ugalj koji se pretvara u öećer i ugljeno hidrate.
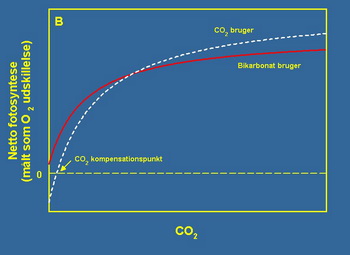
Vodene biljke uglavnom imaju pristup na dva izvora neorganskog ugljenika: ugljen dioksid (CO2) i bikarbonat (HCO3-). Većina vodenih biljaka viöe vole CO2 naspram bikarbonata, jer to moûe da iskoristi iz ambijenta bez ulaganja energije, pored ovoga većina podvodnih biljaka nije u mogućnosti da direktno koristi bikarbonat u procesu fotosinteze. Uopöteno koriötenje CO2 jedne vodene biljke öematski je prikazano na slici 2B. Na grafikonu moûemo definirati CO2 tačku gde koncentracija CO2 ispod prikazane tačke daje rezultat u negativnom delu fotosinteze dok koncentracijama iznad ove tačke daje rezultat u pozitivnom delu fotosinteze. Ovaj grafikon slikovito prikazuje iskoristljivost bikarbonata, odnosno podvodne biljke mogu nastaviti sa procesom fotosinteze u pozitivnom delu čak i kada je koncentracija CO2 na nuli, iz razloga öto koristite bikarbonat kao izvor neorganskog ugljenika.
Slike 2A i 2B. Na grafikonima su prikazane teorijske zavisnosti fotosinteze od svetla (grafikon A) i fotosinteze od koncentracije CO2 (grafikon B ). U oba slučaja zavisnost je opisana saturacionom krivom (krivom zasićenja), iako se oblik samih krivih razlikuje na grafiku A i B tip krivih je isti.
U prirodi, koncentracija CO2 je često veći u vodi nego li u vazduhu (u vazduhu ga ima oko 0,03% ), ali uprkos tome stvarna dostupnost za vodena biljke je niûa. To je upravo zbog sporog kretanja gasova u vodi u kojoj je difuzija (rasprostranjenost) oko 10.000 puta manja u vodi nego u vazduhu. Dakle, iako je koncentracija CO2 u mnogim vodenim povröinama znatno veća nego li u vazduhu, upravo iz činjenice da se gasovi sporije kreću u vodi na kraju dovodi do ograničenja iskoristljive koncentracije CO2 za vodene biljke.
Evolucija je razvila tanke listove koji su uglavnom tipični za vodene biljke, u velikoj meri olaköavaju ovo ograničenja sporijeg kretanja CO2 u vodi. Sama građa listova sadrûi tanje granične slojeve kroz koje se CO2 laköe apsorbuje, kada jednom CO2 prodre u list ne mora dugo putovati pre nego öto veûe ugljenohidrate u procesu fotosinteze. Mora se spomenuti i velika vaûnost, sposobnosti podvodnih biljaka da na gore ili na dole reguliöu različite grupe enzima (FX Rubisco i Pepcarboxylase) koji učestvuju u stabilizaciji ugljenika. Na niskom nivou koncentracije CO2 koji je dostupan, biljka mora da uloûiti viöe energije u enzime koji će joj pomoći u procesu unoöenja CO2 ili stabilizaciji ugljenika i time ublaûiti efekat ograničenja.
Pored ovoga neke biljke su takođe u stanju proizvesti izoenzime, koji su u stvari enzimi različitih hemiskih optimuma tako da sklonost za CO2 moûe da se promeni u korist unoöenja CO2. Joö uvek u potpunosti nije jasno koliko je vaûan izoenzim u procesu CO2 iskoriötenja, u naučnoj literaturi tekstovi vezani za izoenzime su često povezana sa temperaturnim aklimatizacijama unutar samih biljaka.
Adaptacija biljaka na ograničene resurse je momenat kada se javljaju propratni efekti odnosno posledice. Ukoliko biljka poveća formiranje hlorofila ili formiranje viöe enzima kao rezultat dobijamo povećanje za hranljivim elementima. Veća upotreba energije dolazi iz činjenice da proteini zahtevaju stalno odrûavanje na ćeliskom nivou da bi ispravno funkcionirali i ti procesi odrûavanja apsorbovali dragocenu energiju i ugljeno hidrate, koji bi inače mogli biti koriöteni za rast i razvoj biljke.
Ono öto je bilo bitno prlikom proučavanja jeste samo određivanje koji faktor CO2 ili svetlo ima veću vaûnost ili ograničavajući faktor na rast i razvoj vodenih biljaka. Pored ovoga zanimljiv je odgovor na pitanje da li CO2 ili svetlo moûe delovati kao zamena jedno za drugo, a samim time i povećati rast FX.
Da li povećati koncentraciju CO2 pod jakim izvorom svetla? ili ograničenim izvorom svetla?
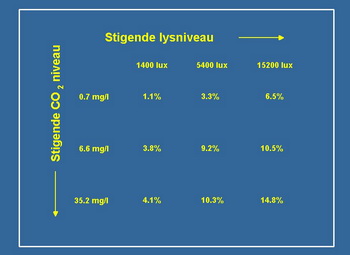
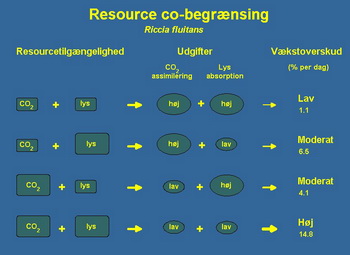
Da biste mogli odgovorili na ovo pitanje eksperimentalno je koriötena Riccia fluitans u labaratoriski kontrolisanim uslovima sa mogućnuöću kontrolisanja nutrijenata (pod nutrijentima se podrazumevaju azot, fosfor i svi ostali mikro elementi). Osmislili smo eksperiment na način da moûemo da reguliöemo intezitet svetla i koncentraciju CO2 tako da sve ovo bude neovisno. U tablica 1 je grafički prikazan eksperiment gde smo imali ukupno devet različitih tretmana razlika je u intezitetu svetla i koncentraciji CO2. Slabiji intezitet svetlost odgovara uslovima koji se mogu naći u prirodi pri samom dnu öto je slično većini standardno opremljenih akvarijuma. Jači intezitet svjetlosti odgovara uslovima koji se mogu naći u prirodi pri povröini vode ili u izrazito dobro osvetljenom akvarijumu sa ûivinim sijalicama visokog pritiska ili halogenim lampama. Srednji intenzitet svetla u ovom istraûivanju odgovara standardu dobro osvetljenih biljnih akvarijuma.
Niska koncentracja CO2 odgovara uslovima koji se mogu naći u prirodi u mnogim jezerima ili u akvarijumu bez opreme za CO2. Visoka koncentracija CO2 je 40 mg / l ovo odgovara uslovima koji se mogu naći u prirodi u malim podzemnim potocima, ujedno je i maksimalna koncentracija koju preporučuju većina iskusnih biljnih akvarista.
U ovom eksperimentu smo se odlučili za Riccia fluitans uglavnom zato öto je ovo biljka koja lako raste i pored ovoga izuzetno je prilagodljiva uslovima. Dakle, moûemo eksperimentalno proizvesti veliki broj ponavljanja, koji nam kasnije pomaûu kod dobijanja rezultata i samim tim izvlačenja zaključka iz naöih eksperimenata.
Tabela 1 prikazuje rezultate tih eksperimenata, u njoj je izraûena dnevna stopa rasta Riccia fluitans, pod pretpostavkom da eksponencijalni rast tokom perioda (1). Moûemo primetiti da je pri slabom svetlu i niskom CO2 Riccia fluitans jedva bila u stanju da zadrûi pozitivan rast, dok na niskoj koncentraciji CO2 i jačem svetlu stopa rasta je gotovo öest puta veća. Joö vaûnije, na niûem intezitetu svetla dodavanjem CO2 uspeli smo stimulirati rast Riccia fluitans faktorom četiri.
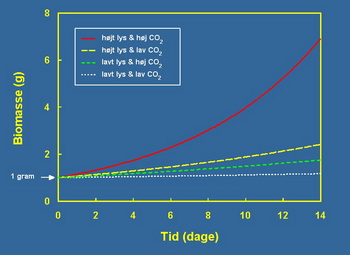
Moûda ovakva stimulacija rasta sa faktorom 4 do 6 ne izgleda puno, ali zbog eksponencijalne prirodne stope rasta to u stvari čini veliku razliku u odnosu na period od, na primer, dve nedelje. Slika 3 pokazuje kako se 1gram Riccia fluitans razvija u periodu od preko dve nedelje sa četiri različite stope rasta.
Slaba svetlost i nizak CO2 jedva rezultira pozitivnom stopom rasta tokom dve nedelje, dok je tretman sa većom CO2 koncentracijom i slabijim svetlom gotovo udvostručio masu Riccia fluitans.
Primera radi ako uporedimo, jače svetlo i niûi CO2 daje kao rezultat povećanje mase od 2,5 grama nakon dve nedelje. Nepotrebno je reći da je korist od povećanja i svjetlost i CO2 nadmaöila rezultat podizanja za samo jedan parametar.
Tabela 1. U tabeli su prikazani eksperimentalni podatci procentualnog dnevnog prirasta biomase u zavisnosti od količine svetla i koncentracije CO2. Pregledanje tabele sa leva na desno odgovara uslovima sa povećanjem količine svetla, dok pregledanje odozgo na dole odgovara uslovima sa povećanjem koncentracije CO2 u vodi. Prva kolona odgovara uslovima slabog osvetljenja (Low light) koja je tako česta u mnogim standardnim akvarijumima, dok prvi red odgovara uslovima koji se takodje često sreću u akvarijumima u kojima rade vazduöne pumpe i kada je akvarijumska voda zasićena vazduhom. Za poredjenje u severnoj Evropi u toku sunčanog dana jačina sunčevog svetla je oko 70000 Lux-a, a koncentracija CO2 u malim potocima moûe da dostigne i vrednosti od 50mg/lit. Jedinice Lux i mg/lit su zastarele jedinice i u savremenoj nauci se koriste druge jedinice. Za svetlost talasne duûine od 400 do 700 nm obično se koristi jedinica 1µmol fotona/ms koji je jednak 60.6Lux, a za koncentraciju CO2 1 mmol/lit koji je jednak 44mg/lit.
Na najviöem intezitetu svetla i CO2 koncentraciji, 1 gram Riccia fluitans dobija na uvećanju mase na neverovatnih 6,9 grama nakon dve nedelje. Iznenađujuće stimulacija promatrana u odnosu na stopu rasta kada su svetla i CO2 povećani je veća od dodatog doprinosa svakog pojedinog parametra.
Na primer: stopa rasta sa slabim osvetljenjem i niskim CO2 je 1,1% po danu. Povećanjem svetla, biljka raste 3,3% po danu ili dodatnih 2,2% u odnosu na početne uslove. Isto tako, povećanjem CO2 stopa rasta je povećana 3,8% na dan ili dodatnih 2,7%. Iz toga izvodimo zaključak da je doprinos povećanja 6,0% po danu (1.1 + 2.2 + 2.7), ali rezultat stope rasta sa kombinacijom svetla i CO2 je 9,2% po danu, öto je znatno veći.
Uzimajući u obzir eksponencijalni rast moûemo koristiti sledeću formulu za izračunavanje stope rasta: μ = (W1 - W0) / t, gdje je W0 teûina biljnog tkiva na početku eksperimenta, W1 je teûina nakon inkubacije i t je vreme inkubacije izraûeno u danima. Osnova za pretpostavku eksponencijalnog rast je činjenica da se svo novo tkivo formiralo tokom inkubacije i nastavlja sa daljim formiranjem u daljem toku same inkubacije.
Grafikon 4 prikazuje konceptno objaönjenje naöih nalaza. Prlikom slabog osvetljenja i niske koncentracije CO2 nema puno energije sa kojom se moûe raditi u smislu povećanja ili sniûavanja zaliha hlorofila ili enzima. Ali smo radi ekeperimenta povećali koncentraciju CO2 u sistemu biljke, kao rezultat dobili smo manje ulaganja energije i resursa za apsorbovanje CO2 sami tim ostaje viöe energije za optimalnije iskoriötavanje svetla - viöe hlorofila moûe biti proizvedeno bez fatalnih posljedica za iskoriötenje energije do maksimuma. Iz toga proizilazi, iako nije podignut intezitet svetla, biljka sada moûe koristiti raspoloûivo svetlo daleko efikasnije. Potpuno isto objaönjenje se moûe koristiti da bi se objasnilo kako povećanje svetlost moûe podstaknuti rast čak i pri vrlo niskim koncentracijama CO2.
Slika 3. Na grafikonu je prikazan prirast (vertikalna osa) jednog grama Riccie u toku vremena od dve nedelje (horizontalna osa) i u zavisnosti od količine svetlosti i CO2 u koji su dostupni biljci za rast. U uslovima slabe svetlosti i niske koncentracije CO2 Riccia jedva uspeva da zadrûi biomasu i 1 gram se za 2 nedelje uvećao na 1,16 grama (bela krivulja). U uslovima slabog svetla i velike koncentracije CO2 od jednog grama Riccie posle 2 nedelje biomasa je porasla na 1,75g (zelena kriva), u uslovima jakog svetla i niske koncentracije CO2 na kraju smo imali 2,41g Riccie (ûuta kriva). Kombinacija jakog svetla i velike koncentracije CO2 prevazilaze efekat koji se dobija prostim sabiranjem krivih kada je samo jedan od resursa u viöku (Co2 ili svetlo). U uslovima jakog svetla i puno CO2 za 2 nedelje biomasa Riccie je narasla sa 1g na 6,90g.
Slika 4. Diagram simbolično prikazuje kao na prirast Riccie (treća kolona) utiče raspoloûivost resursa (CO2 i svetlo) u okruûenju (prva kolona) i kolika energije biljka troöi (srednja kolona) u zavisnosti od raspoloûivih resursa. Jednostavno rečeno: ako je dostupnost resursa (svetlo i CO2) u spoljaönjoj sredini mala biljka troöi puno energije na njihovo ìhvatanjeî, a prirast je jako mali, öto se vidi u prvom redu tabele. Poslednji red odgovara uslovima sa obiljem resursa u okruûenju, pa biljka ne mora da troöi puno energije na njihovo usvajanje, a prirast bio mase je ogroman.
Verujemo da rezultati ovog eksperimenta koji su rađeni na Riccia fluitans mogu biti primenjeni na većini vodenih biljaka. U posljednjoj deceniji je doneöeno viöe naučnih teorija i dokaza koji podrûavaju ove rezultate. Tu su eksperimenti sa Elodea canadensis i Callitriche sp. Oni pokazuju istu tendenciju (pogledajte spisak literature) ukazuju na to da ograničenje resursa nije tako jednostavno kao öto je Justus Liebig opisao u svom radu.
Mnogi izvori su u stanju da zamene jedni druge ili barem da ublaûe simptome ograničenja resursa.
Sagledavanjem tih podataka sa viöeg globalnog aspekta, moûemo očekivati da će u budućnosti povećanje koncentracije CO2 u atmosferi zapravo imati kao rezultat da se moûe dovesti do povećanja biljne proizvodnje na zemlji.
Međutim, također moûemo predvideti ozbiljne sporedne efekte. Biljke koje rastu na poviöenim koncentracijama CO2 moglu da razblaûe svoje zalihe vezanog ugljika za enzime i da će samim tim smanjiti vrednost proizvodnje u vidu enzima i proteina. Za vodene biljke, udvostručenje koncentracije CO2 u atmosferi verovatno neće imati nikakvog uticaja na biljnu proizvodnju, jer većina vodenih biljaka već rastu pod prezasićenim CO2 uslovima. Za one vrste biljaka koje rastu periodično u emerznoj odnosno periodično u submerznoj formi, teöko je predvideti uticaj na povećanu dostupnosti CO2, jer ovde moramo uzeti u obzir i uticaj fitoplanktona.
Neko se moûe zapitati kako moûemo koristiti sve gore navedene informacije u samom hobiju biljnih akvarijuma! Na mnoge načine, moderna oprema akvarijuma podseća na naöu eksperimentalnu postavku sa Riccia fluitans. Iako su svi pojedinačni resursi teöki za savröeno kontrolisanje, mi smo u stanju odrediti koliko svetla, koliko CO2 i koliko hranjive materije u obliku azota, fosfora, gvoûđa i mikroelemenata moûemo da ponudimo naöim biljkama. Počevöi sa hranjivim materijama, prosečna akvarijumska biljka sa normalnom populacijom riba obično ima dovoljno azota i fosfor. Kada je u pitanju gvoûđe, kalijum, mangan i drugi mikroelementi tu su već stvar malo teûe.
Neki akvaristi već u startu dobro planiraju od samog početka formiranja akvarijuma, na primer laterit i druga đubriva u podlozi, dok drugi to ne uzimaju u obzir. U većini slučajeva, akvarijumske biljke bez azota fosfora i đubriva sigurno mogu biti odrûavane na nivou zdravog rasta. Međutim često je mnogo teûi i zahtevniji zadatak osigurati odgovarajući izvor svetlosti iznad biljnog akvarijuma. Fluoroscentne cevi i sijalice pod visokim pritiskom mogu proizvesti dovoljan intezitet svetlosti ukoliko su isporučeni sa kvalitetnim reflektorima, ali u dubokom akvarijumu (preko 50 cm dubine), vrlo je teöko ponuditi dovoljno svetla za niske biljke prvog plana koje su obično najzahtevnije po pitanju inteziteta svetla.
Na temelju naöih eksperimenata, predlaûemo dodavanje CO2 prije bilo koje druge radnje! Verujemo da je čak i sa vrlo skromnim intenzitetom osvetljenja moguće doûiveti primetne promene u biljnoj masi unutar vaöeg akvarijuma. Oko tačnog iznosa koncentracije CO2 uvek se moûe voditi polemika, ali ako nemate jako osetljive ribe, odrûavanjem koncentracije CO2 od 25 mg do 50 mg / l to će samo da poboljöa rast biljaka. Vervatno ćete primetiti da će biljke, koje su jedva bile u stanju da preûive prije povećanja koncentracije CO2 sada bolje napredovati u prisutnosti veće koncentracije CO2.
Koriötena Literatura:
Andersen (1999) Interakcija između svetla i neorganskog ugljenika stimuliöe rast Riccia fluitans L. Izvjeötaj iz Slatkovodne bioloöke labaratorije, Univerzitet iz Kopenhagena (e-mail tandersen@zi.ku.dk), na Danskom.
Maberly (1983) Međuzavisnost fotonskog zračenja i slobodnog ugljendioksid ili bikarbonatne koncentracije na fotosintetičku nadoknadu slatkovodnih biljaka.
Maberly (1985) Fotosinteza i Interakcija između fotona zračenja, koncentracije ugljendioksida i temperature.
Madsen (1993) Rast i fotosintetska aklimatizacija po Ranunculus aquatilis L. odgovor na neorganski dostupan ugljenik.
Madsen i Pijesak-Jensen (1994) interaktivni uticaj svetla i neorganskog ugljenikana na rast i razvoj vodenih biljaka.